From W. Reich’s book, Cosmic Superimposition: Man’s Orgonotic Roots in Nature, Orgone Institute Press, pp. 27-52
The formation of living matter in orgone Experiment XX[1] [2] combines numerous bio-energetic and biofunctional phenomena into a single result of great significance. This experiment reproduces the process of primary biogenesis, i.e., the original formation of plasmatic, living matter through condensation of mass-free cosmic orgone energy. This conclusion derives logically from the fact that organic forms with all the properties of living matter (structure, pulsation, reproduction, growth, and development) can be developed by a freezing process in a clear solution of high orgonotic potency. The subject is inexhaustible, but it is not our objective to treat it exhaustively. Once more, I would recall the discovery of the American continent by Columbus. This discovery did not exhaust all past and future possibilities of America. It did open the door to an enormous territory full of future potentialities. The same holds true for Experiment XX.
The schema below represents approaches to the manifold functions of nature that were opened up by Experiment XX:

- The development of organic forms, plasmatic “orgonomes” (bions).
- The organization of protozoa (orgonomia).
- The formation of biochemical matter: coal, sugar, fat.
- The furtherance of life and growth as affected by the orgone-water solution.
In the process of the freezing experiment, energy is transformed into matter. This matter is alive. By means of dehydration or burning of the flakes, carbon and a sweet-tasting, sugary substance originate from it. These are gross characteristics to be elaborated in detail. In this process, frozen orgone energy passes through all phases of bionous formation revealed by orgone biophysics: T-forms develop into PA bions through the intake of mass-free orgone energy; the PA bions grow into larger, rounded shapes resembling small “eggs”; some of these “egg shapes” expand and become beanshaped; the bean shapes acquire motility and form protozoa: orgonomia. In movement and shape, they look very much like spermatozoa. We may assume that the spermatozoa and eggs in the metazoa are also formed through condensation of orgone energy in the germinal tissues. The development of formed bions from distilled orgone water establishes beyond any doubt the process of primary formation of organic matter from mass-free orgone.
Bion water is yellow, ranging in intensity to almost a brown. In this context, one is reminded of the yellow resin produced by trees, of yellow honey produced by bees, of the yellow color of animal blood serum, the yellow of urine, etc. Also of great significance is the “blood sugar level” in the organism. Thus the gap in biology, which up to now contained a mystery—namely, how plants convert “solar energy” into carbohydrates and solid cellulose forms—is apparently closed. “Solar energy” is our orgone energy that the plants absorb directly from the soil, the atmosphere, and the rays of the sun.
The leaves of evergreen ivy are a case in point. In winter, the leaves lose their green color, except for the venation, which remains green. The rest turns a yellow-brown. In spring, the green expands from the leaf vessels across the smooth leaf. This phenomenon permits the assumption that, in winter orgone energy retreats from the periphery of the leaves; in other words, it contracts because of the cold, to expand again in spring. That portion of the ivy leaves about to die off is thus revived.
The change from green to yellow in autumn and from yellow to green in spring becomes perfectly comprehensible in terms of orgonotic functioning. According to classical investigations, green is the result of a mixture of yellow and blue. Blue is the specific color of orgone energy, visible in the atmosphere, ocean, thunderclouds, “red” blood cells, protozoa, etc., and on orthochromatic photographic plates after irradiation with earth bions.
Now it seems clear that the yellowing of the leaves in autumn is due to disappearance of the blue from the green, and accordingly, the turning toward green again in evergreen ivy is due to new absorption of orgone energy from the atmosphere. Thus, the green of leaves is the result of the mixture of yellow resin and blue atmospheric orgone energy.
At present, we would like to limit our investigation to a single function: the origin of formed living matter from mass-free orgone energy. Right now we are not interested in the chemical composition of these forms.
There is only one assumption that satisfactorily explains the origin of motile, formed living substance in Experiment XX. In the process of freezing, the mass-free orgone energy in the fluid contracts, just like living plasma. Hence, this contraction does not depend on the existence of formed matter. It exists prior to the formation of matter, as a basic function of cosmic orgone. The contraction of orgone energy is accompanied by condensation, and condensation is accompanied by the formation of material particles of microscopically small dimension. The classical, mechanistic concept does not provide for any causal connection between energy movement and organismic form. Orgone biophysics can prove a functional connection between form of movement and form of living matter.
Primary matter originated in the cosmos, and the process of matter formation apparently continues uninterruptedly. The cosmic origin of bio-energy is experienced as an equation of “life-earth-sunspring.” The mechanistic concept knows only atoms and molecules that combine to form salts and organic bodies. It can explain neither movement nor formation of living matter because neither the first nor the second resemble mechanical movements and known geometric forms in any way. In contrast, orgone biophysics operates with a concrete cosmic energy. It postulates that the functions of cosmic energy in the realm of inorganic matter are in harmony with those in the realm of living matter.
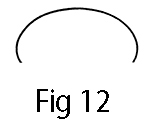
In Experiment XX, membranes and then bions are formed from mass-free orgone energy. They constitute forms that cannot yet be described as “living organisms” in the accepted biological sense, but they already show the typical shape of living organisms. This is clearly apparent in the illustrations of Experiment XX (cf. fig. 19). The forms of most flakes resemble those of fish or tadpoles. Now, if forms invariably express frozen movement, we may reason, a posteriori, from these forms to the functions of orgone energy. Exact observation and extensive comparison will show that there exists a basic form of living matter that has no counterpart in classic geometry. Viewed laterally, this basic form looks as follows:

Viewed from above or below, the living form is typically as follows:

Before investigating the bio-energetic function of this form, let us make sure that it is indeed the biophysical basic form. It clearly applies to:
- Plant seeds: wheat, corn, barley, oats, beans, lentils.
- Plant bulbs: potato tubers, almond kernels; the pits of apples, pears, plums, peaches.
- Animal sperm cells.
- Animal eggs, particularly birds’ eggs.
- Animal embryos.
- All organs of the animal body: heart, bladder, liver, kidney, spleen, lung, brain, testicle, ovary, uterus, stomach.
- Unicellular organisms: paramecia, colpidia, vorticellae, cancer cells, human vaginal protozoa (trichomonas vaginalis) etc.
- Whole animal and plant bodies: jellyfish, starfish, reptiles of all kinds; the trunk formation of all kinds of birds, fish, beetles; mammals, including man, etc.
- Trees in general, as well as each single leaf and blossom; pollen and pistils of plants.
It is noteworthy that even those organs extending from the trunk—arms, legs, fins, wings, the head of the snake, the lizard, the fox, man himself, the fish, etc.—in turn take the form of the “orgonome.” Even the claws and beaks of birds, the air bladder of fish, the horns of cattle, rams and stags, the shells of snails and mussels take the form of the “orgonome.”
All this points to the work of a natural law of energy, a law that fundamentally differs from the geometric laws of classic mechanistics.
Access to this law of cosmic energy must be sought in the movement of mass-free orgone energy.
Just as the expressive movements of living matter are inextricably tied to an emotional expression that is meaningful in relation to the world around it, so the form of living matter has a specific expression too. The point is to read it correctly.
All forms in the realm of living matter can easily be reduced to the egg form without violating the individual variations of form. This basic form varies with length, width and thickness. It may appear in subdivisions of the same form, as in worms; but whether as a whole or in part, the basic form of living matter always remains the same egg form.
Such a consistent uniformity of the organic form must correspond to a fundamental law of nature and a natural law of cosmic dimensions. For the basic biological form is universal, regardless of climate or geology. It is as if cosmic orgone energy, in organizing living substance, obeyed only one law, its own law of motion.
We shall call the specific basic form of living matter the orgonome. Its typical basic form is the following generalization of microscopic forms from Experiment XX:

Trigonometry of the orgonome
We would like to designate as an orgonome that specific form which, in its purest shape, is represented by the hen egg.
The orgonome is neither a triangle nor a square nor a circle; it is neither an ellipse nor a parabola nor a hyperbola. The orgonome represents a special, novel geometric figure, a closed plane curve, not unlike the ellipse with half-axes of varying length and width, but differing from the ellipse precisely because of the different length of the large axes.
Let us try to determine how an orgonome originates, orgonometrically speaking. Two fundamental natural phenomena are involved:
- The orgastic convulsion.
- The spinning wave—Kreiselwelle (KRW).
We encounter the orgastic convulsion in the entire animal kingdom. We discern the spinning wave (KRW) by observing atmospheric orgone in the darkroom. The tiny, blue-violet points glide along specific trajectories, which I described schematically in the second volume of The Discovery of the Orgone as follows:

Let us isolate an individual wave from the KRW path:
If we place two such spinning waves together with their concave sides, we obtain the known form of the ellipse:
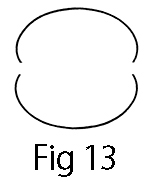
However, if we bend a KRW in the center at point A and bring the two ends of the KRW—B and B’ —together, we obtain the egg, or orgonome form.

We could work out this process in terms of pure trigonometry without providing orgone-physical proof. But the orgastic convulsion gives us a biophysical argument that endows this trigonometric process with great importance. The most conspicuous phenomenon in the orgasm reflex is the striving of both ends of the torso—the mouth and the genitals—to come closer together. This biophysical phenomenon indeed put me on the trail leading to the origin of the orgonome form. In the orgastic convulsion of an animal, or in the swimming motion of the jellyfish, the body seems to sag in the center, bringing both ends closer together.
The connection of a fundamental biological movement with a physical movement may, at first glance, seem arbitrary. But such a connection is justified if it opens the door to an obvious lawfulness in biological functioning. To my knowledge, the basic form of living bodies has never been understood. And if the orgasm reflex promises us an understanding of the orgonome form, we must not reject it.
The similarity of a KRW to an animal body, viewed laterally, is indeed startling (cf. fig. 14). Detailed proof of this similarity cannot be presented here but has already been established experimentally.
If living matter is frozen orgone energy, the form of movement of the orgone energy must necessarily translate itself into the form of living matter, the orgonome form. This functional continuity is hard to find in the realm of inorganic matter. It is easily understandable in the realm of living matter. If form is the movement of energy that is frozen, then the organ form must derive from the form of movement of cosmic energy.
Let us return to the orgasm reflex, this rich source of bio-energetic insights:
We found that the orgasm reflex cannot be verbalized in terms of idiomatic language. Its mode of expression, we concluded, was supraindividual—neither metaphysical nor mystical, but cosmic. In the orgasm reflex, the highly excited organism attempts to bring both ends of its torso closer together as if to unite them. If this interpretation is correct, it must also prove correct in other categories of orgone functioning and cannot be limited to the orgasm reflex alone.
Let us now look at the form of the biological orgonome in its functional connection with the form of plasmatic currents. True to the principle of the functional identity of all living substance, we must gather apparently widely separated functions and look for their common denominator.
Plasmatic current does not flow continuously but in rhythmic thrusts. Hence we speak of pulsation. The pulsation can be plainly observed in the blood circulation of all metazoa. The pulsatory current of body fluids is the work of the organismic orgone, a direct expression of its form of movement. From the pulsation of body fluids we must reason, a posteriori, that there is a pulsation of orgone energy. This conclusion is confirmed by observing certain protozoa, in which pulsatory waves of excitation pass through the body and set the protoplasm in motion. Among worms, excitation waves of a pulsatory nature pass from the tail end to the head. The same phenomenon can be seen in certain amoeboid cancer cells. The following drawing expresses the form in which excitation waves move in the protoplasm of these cancer cells:

Hence we must distinguish between two kinds of pulsatory movements in living matter: the pulsatory movement of orgone energy in the organism, and its effect, the pulsatory mechanical movement of body fluids. We differentiate here between functional bio-energetic pulsation and mechanical pulsation. The mechanical pulsation results from the functional pulsation of the orgone, its spinning forward in alternating expansion and contraction.
Since the movement of fluids is mechanical, it can only be the expression and consequence of the pulsatory function of orgone energy. Among the flowing amoebae, the bio-energetic pulsation coincides completely with the organic flow of fluids. Among the colpidia and paramecia, the body is rigid and contains large, membranous fluid-filled vesicles without flowing plasm. Here the movement of energy can be discerned only in the locomotion of the whole body. If we compare the form of movement of the waves of excitation in cancer cells with the external form of movement of trichomonas vaginalis, colpidia, and paramecia, we find there is a thrusting, pulsatory motion that does not proceed in a straight line but in the manner of a spiral, presenting an overall curvature. We can connect the individual points of the movement curve and find a geometric figure that depicts a spinning wave (KRW) and looks roughly as follows:

We see that the curve of the plasma current inside the body of the cancer cell is the same as in the locomotion of the whole body of a colpidium. If we dissect the curve of the orgonotic plasma current into its individual parts, we obtain a shape that, laterally viewed, resembles the form of all living organs and organisms (cf. fig. 14).
This harmony, in the form of movement of the energy particles, plasma current, orgonotic excitation waves and the shape of the organs, cannot be mere coincidence. It is obviously governed by a common law of movement revealed time and again in the individual forms of motions and structures. Even the elongated earthworm, which, at first glance, reveals nothing that resembles an orgonome form turning back on itself, shows the orgonome in the segments. Furthermore, the earthworm curls up in a manner that looks like the orgonome of a snail shell (cf. fig. 20:3 and 4).
The following diagram illustrates the structuralized, clearly expressed original movement of the organismic orgone energy in the growth of a shell:
Thus we can distinguish three states of orgonotic expressive movement:
- The spinning motion of orgonotic excitation waves, of protoplasm and of the locomotion of protozoa.
- The orgonome form of animal organs and organisms, i.e., frozen orgone movement.
- The orgonome form of the animal body at rest, as an intermediary state between energy movement and solid matter.
We now have a better biophysical understanding of the segmentary arrangement of the orgonotic current in man and of the segmentary arrangement, or armoring, in the biopathic character.
The plasmatic (mechanical) and the orgonotic (bio-energetic) currents in man—blood circulation and excitation waves—have the same rhythmic, wavy, and segmentary character as observed in the earthworm. The segmental arrangement of the armoring expresses the immobilization of individual parts of the wave path, or, to put it differently, one wave freezes into one formed orgonome segment.


Thus the principle of orgone therapy—to proceed always from the “head” to the “tail,” i.e., to the genitals—acquires its bio-energetic meaning. As in the earthworm, the snake and the plasmatic cancer cell, the orgonotic waves invariably pass from the tail end over the back toward the head. Bioenergetically, this arrangement of the orgonotic flow makes sense, because it is predicated on the “forward” movement of the whole body in the direction of the head. In orgone therapy, if we first loosened the armoring at the tail end, the liberated energy would be blocked at the segment located farther ahead. But the dissolving of the armor at the head end eliminates the armor rings at the place toward which the orgonotic excitation must flow. We meet the direction of the current, and thus free the way for its unhindered flow, instead of starting to break the armor at the source of this current. The technique of orgone therapy did not start out with these biophysical speculations in mind, but followed purely clinical considerations, e.g., that it would be advantageous to liberate all the energy of the body before mobilizing the genitals. But, as we now see, the clinical and the bio-energetic aspects of the matter combine in a common useful function.
Let us now return to our Experiment XX in order to learn more about the formation of living substance into the orgonome. We find plasmatic flakes in which first circular, then bean-shaped orgonome forms can be seen. In the bean shape, the orgonome is once more clearly evident. This orgonome is in motion. Its movements again have orgonome form, as can easily be discerned in the spiral lines of their progressive movement.
We may now appropriately conclude that through the freezing process the freely moving orgone energy in the fluid is, in very small part, converted into matter by membrane formation. Since the movement of orgone energy is curved, it stands to reason that the membranes are also curved. Inside the membranes, mass-free orgone energy continues to move. Naturally enough, it strives to expand the membrane, as if it meant to burst through the sac in which it is trapped. There is of course no reasoning involved here, but rather a contradiction between the function of the expanding movement of the mass-free orgone and the confining membrane. Logical deduction demonstrates that nothing but a bean shape, our orgonome, can result from this contradiction between energy flow and restricting membrane.
Of course, the formation of the bean shape does not in any way satisfy the motile impulse of mass- free orgone energy inside, an impulse directed toward stretching the curve, i.e., toward moving away from the spot. Therefore, the local forward movement, whose basic tendency consists again in stretching, curving, and rhythmically reverting upon itself, appears for the first time.
The development of colpidia from primary embryonic vesicles is particularly suited for studying the plasmatic currents that are set in motion by the orgone energy in the membranous sac. As soon as a membrane has formed around a cluster of bions, the budding germinal vesicle appears. The interior shows a vesicular structure and a blue glimmer. The membrane is taut, but the whole system is still at rest (fig. 21: 1). That motile impulses are freed in the interior of the “germinal vesicle” is shown by a rolling motion of the vesicles occurring sooner or later. While the membrane rests, the vesicles at first roll near the periphery, in one direction along the membrane. The inner cohesion loosens. Along with the rolling motion in one direction goes a reciprocal attraction and repulsion. After a while, the direction of the movement changes; the vesicular content reverses its direction. In this manner, the bionous content gains elasticity (21: 2). The germinal vesicle tautens more and more; it grows larger. Gradually the circular form turns into the egg form, our orgonome form. The plasmatic current at one end splits into two currents. The two currents converge and continue backward along the center line (21: 3). Now we can clearly distinguish two halves of the orgonome, each of which assumes more and more clearly the bean shape, or lateral orgonome form. After several hours of strong orgonotic motility of the plasm, the germinal vesicle usually bursts into four “complete” colpidia. So far, we cannot determine whether the figure “four” is the rule or whether a division into two colpidia also occurs. What is important is that the forward end of the colpidium is located at the place where the current was originally directed. The animalcule swims off locally in the direction of the original plasma current (21: 4). This current has assumed an orgonome form. Now, when the local movement begins, the internal current stops and the animal moves forward as a whole in lines that are slightly curved. The curve of the path of locomotion is identical with the curve of the “back,” as illustrated in drawings from life (fig. 21).

Let us summarize the processes in the living orgonome.
- The inner motility is nourished by wave-like pulsating orgone energy that is trapped in a membranous “sac.”
- The movement of the orgone energy is responsible for the inner motility of the structured bionous substance.
- Since the inner orgone movement is confined by the membrane, a curved path of the plasma current is produced, in which we recognize the orgonome.
- The “energetic” orgonome leads to the formation of the material orgonome. The form of the organs reflects the form of the original energy movement.
- There is a contradiction between the movement of the orgone energy and the taut membrane. The membrane sharply deflects the original forward movement of the current backward. Since this happens at all the curvatures of the vesicle, the currents converge toward the center and thus produce a division of the vesicle into four structural orgonomes.
- Once this division is complete, we observe the separation and local forward movement of the individual orgonomes. The local movement proceeds in a curved line—a motion with alternating long and short half-waves. The motion “away from the spot” is obviously dictated by the direction of the orgonotic impulses. It is curved in terms of the “back.” The fore end is always located in the direction of the original orgonotic current.
Orgonotic Superimposition
To summarize: The specific orgonome form of living matter and its organs results from an opposition between mass-free orgone energy and frozen orgone that has become membranous matter. Mass-free orgone always strives to break beyond the enclosure of the membrane. The bio-energetic orgonome is extended and open; the material orgonome is closed. Since the excitation waves of the bio-energetic orgonome move within the limits of the closed material orgonome, they necessarily press against the membranous boundary, as shown in the following drawing:

This creates a stretching of the orgonome, in which we recognize the basis for all kinds of growth, particularly as shown in the stretching of the gastrula as it becomes the typical elongated embryo of a multicellular organism (metazoan).
The function of growth corresponds to the expansion of the membranes of the closed orgonome. That this indeed involves expansive functions of mass-free orgone energy can be seen from the curved protrusions that initiate the formation of every new organ in the embryo of all animal species. Again, the embryonic protrusions show the typical form of the orgonome.
The elasticity of the formed body membrane and the presence or absence of a skeleton determine how much of the original spinning motion of the bio-energetic orgonome is clearly evident. But even where a fully developed skeleton and an extensive muscular structure have blotted out the external appearance of the excitation waves, there is still the rhythmic excitation and current pulse of the blood circulation, as well as the orgonotic current or plasma excitation, which are felt subjectively. In the orgasm reflex the original form of movement of the bio-energetic orgonome is unmistakably perceived insofar as it seizes the entire organism.
We distinguish the following kinds of superimposition:

The superimposition of two closed orgonomes is the bio-energetic basis for the superimposition of two organisms during copulation (cf. fig. 25). In this process, the highly excited tail ends penetrate each other bodily; the two orgonomes merge bio-energetically to form a single highly charged energy system. It is characteristic for the homogeneousness of all processes in the living realm that the energy functions of excitation, superimposition, interpenetration, and fusion are repeated in the same functions of the reproductive cells. For, during copulation, sperm cell and egg cell continue the function of superimposition and fusion of the male and the female orgonome, although the division of living orgonomes into male and female individuals remains mysterious even from the standpoint of orgone physics.
Let us now try to comprehend the expressive movement of the orgasm reflex on the basis of the orgonome as the fundamental biophysical form of living matter.
It cannot be the function of the orgasm reflex, as one might assume from the purely teleological standpoint, to carry the male semen into the female genital organ. The orgasm reflex occurs independently of the ejaculation of semen, because we also find it in the embryo—in the typical forward position and convulsion of the tail end; in the rocking, bio-energetic forward motion of the tail end of many insects, such as wasps, bees, and bumblebees, as well as in the usual position of the pelvis and the hind legs among dogs, cats, and hoofed animals. These examples should suffice to demonstrate that the orgasm reflex has a far more general life function than mere fertilization. The mechanistic and finalistic interpretations do not work in this area; they are too narrow and do not reach the heart of the matter.
Let us try to interpret the function of the orgasm reflex in terms of its expressive movement.
The living orgonome, be it an embryo, an insect, or a more highly organized animal, is essentially characterized by the following:
First, local forward motion invariably and logically proceeds in the direction of the larger and wider fore end. Second, the genital organs are invariably and logically located on the ventral side near the tail end. Third, in a state of orgonotic excitation of the orgonome, the genital organ expands through erection in the direction of the local forward motion. Fourth, the movements that cause the interpenetration and fusion of the male and the female genital organs drive the entire tail end to the fore in a highly energetic manner (cf. fig. 24).

These biological phenomena are valid for the animal kingdom at large, except for those species that have barely progressed beyond the stage of the primitive orgonome form of the jellyfish. Although they seem to be far apart, there is nevertheless a close functional interconnection. It can be found if we are again guided by the process of orgonotic excitation.
The form and position of vertebrae among vertebrate animals reveal the direction of orgonotic excitation waves during growth: they always start from the tail end and move over the length of the curved back toward the head end. They also follow the same direction during the entire lifetime of the organism. This can be experienced subjectively if shudders of pleasure or fear pass over one’s back. The fur of frightened animals can be seen to “stand on end” due to contraction of the mm. erectores pilorum in the direction of the orgonotic wave motion, leaning forward.
As we can see from the drawing (fig. 24), the entire back is gently curved and as such is in harmony with the curved path of the orgonotic waves. Presumably, the curve of the wave path conditions the curve of the back, and not the other way around. But once the material, closed orgonome is formed, it confines the bio-energetic waves of excitation and forces them to deflect from the original path of the extended course. It is probable that the generally frontward formation of the secondary protrusions during the growth of the embryo is associated with this process. Here, the essential point is the opposition between the material and the bio-energetic orgonome. The membrane of the material orgonome returns from the fore end to the tail end, forming a characteristically wide curvature. In the animal embryo the curvature of the orgonome turns in at the neck toward the body center, then turns away from it near the chest. The curvature of the orgonome forces the excitation waves back toward the tail end. Part of the orgonotic excitation apparently is indeed deflected toward the tail end, but another part pushes through the membrane at the fore end in the direction of the original excitation waves of the bio-energetic orgonome.
As long as the directions of the material and the bio-energetic orgonome are in harmony, there are no new formations and no directions of movement of the whole. The body orgone does not press outward from the orgonome sac. Therefore, no organs are formed along the length of the backs of animals, no protrusions of any kind, but neither is there any movement in the direction of the back, nor any growth. The humps on the back of the camel or the dorsal fins of certain fish are exceptions that remain to be explained.
Growth in the vertical (longitudinal) axis and local forward movement thus appear as functions of the body orgone energy, the result of its attempt to burst through the confining membrane sac. The membrane “goes along,” i.e., it expands and thus forms the protruding sacs of the organs in their primitive condition.
In contrast to the back, where the material and bio-energetic orgonomes are in harmony, we find at the fore end on the ventral side a multitude of organ formations of various kinds: the domed forehead, the nose or snout, chin, breasts, the limbs, and the genitals. Now, if our functional concept of organ formation is generally valid, organs formed by protrusion of membranes must always originate at the ventral side, where the direction of the current of biological energy is deflected from its regular course, i.e., wherever the body orgone “strives to break out of the sac” (cf. fig. 24).
We see from our drawing that the progress of the membrane on the ventral side indeed runs counter to the original and true direction of the orgone waves. Consequently, we find time and again, at almost regular intervals—such as in the arrangement of the limbs and the nipples of the breasts—a rhythmically recurring tendency to break through. This contradiction between membrane and energy wave reaches its culmination at the tail end. The tail end is pointed and sharp; the material orgonome moves sharply forward again in the direction of the forward movement of the waves of excitation.
The strong forward propulsion of the tail end among animals, based on concentrated orgonotic wave excitation pressing outward, explains the “genital excitation” and the orgasm reflex in a satisfactory and probably complete manner. The overwhelming pressure of the orgonotic excitation in the pointed and narrower tail end and especially in the less spacious genital organs is explained by the concentration of orgone waves in a narrow space. The orgone energy, deflected from the head to the tail, i.e., opposite its natural direction, presses toward the genital organ in the original forward direction, exciting it and forcing it forward into erection.
We can now interpret the copulation of animals from a functional bio-energetic, i.e., orgonomic, standpoint. The orgone, pressing forward and concentrated in the genital organ, cannot escape from the membrane. There is only one possibility of flowing out in the intended direction—fusion with a second organism, in such a way that the direction of excitation of the second organism becomes identical with the direction of the orgone waves in the first. This process is actually achieved in orgonotic superimposition as shown by the drawing (fig. 25). We see that, with the superimposition of the two orgonomes and with the interpenetration of the genitals, the pressed and therefore “frustrated” tail end can allow its orgonotic waves of excitation to flow in the natural direction, without having to force them back sharply, and that furthermore the space in which these waves can run their course is widened.

Our assertion, according to which the orgasm reflex has no immediate linguistic meaning, is correct. Its function lies beyond language. Yet it expresses something concrete: superimposition follows orgonotic interpenetration. The preorgastic body movements and especially the orgastic convulsions represent extreme attempts of the mass-free orgone of both organisms to fuse with each other, to reach into each other.
I said earlier that the bio-energetic orgonome always strives beyond the realm of the material orgonome. While the energy of one organism flows into the energy system of the second organism, mass-free orgone energy actually succeeds in transcending the limits of the material orgonome, i.e., the organism, and, by merging with an orgonotic system outside its own, it continues to flow. This takes into account the tendency toward stretching, toward widening the effectual area of mass-free orgone energy. In the acme of excitation, large quantities of energy are indeed flowing out, along with genital substances. This process is connected with the subjective sensation of “release,” “liberation,” or “satisfaction” (“gratification”). Since language directly reflects the function of the bio-energetic process, these words express exactly what happens.
Orgastic longing, which plays such an enormous role in animal life, now appears to express this “striving beyond one’s own self,” this “yearning” to escape from the narrow confines of one’s own organism. Perhaps here lies the answer to the riddle of why the idea of dying is so often represented in the orgasm. In dying, too, the biological energy reaches beyond the confines of the physical sac in which it is imprisoned. Thus the irrational religious concept of “liberating death,” of “salvation in the hereafter,” finds its true basis. The function that in the naturally functioning organism is fulfilled by the orgasm in sexual superimposition appears in the armored organism as the nirvana principle or as the mystical idea of salvation. The religious, armored organism expresses it directly: it wants to “free the soul from the flesh.” The “soul” represents the orgonotic excitation, the “flesh” the surrounding confining tissues. The concept of “sinful flesh” has nothing to do with these facts. It is a defense mechanism in the pornographic structure of the human animal.
In summarizing, we may emphasize the simplicity of functional laws of living nature as one of their main characteristics. Functions as widely separated as growth, locomotion, and genital excitation can be reduced to the common denominator of the relationship between mass-free orgone energy and orgone energy that has become matter. The variations of this functional identity (common functioning principle) result secondarily in terms of the location in which this relationship appears in the organism. The width of the sac and its position (at the fore or rear end) dictates whether the deflection of the orgonotic current is expressed as growth energy or as sexual energy. But, seen functionally, all subsequent functions of living matter originate in the primal contradiction between the material and the bio-energetic orgonome. On the basis of this contradiction in living matter, one is even tempted to trace the connections that form the transition to the “highest” contradictions between “materialistic” and “spiritualistic” philosophy. But such an undertaking transcends the competence of this investigation and must be left to further research.
We shall encounter again the function of orgonotic superimposition in the natural realms of biochemistry and astrophysics. For it is orgonotic superimposition that connects the living organism with nature surrounding it. Living matter arose from inorganic nature as a special variation and, in its superimposition, is functionally identical with it. From here the path leads to the orgonometric investigation of the functional principle of nature per se.
[1] The development of organic forms, plasmatic “orgonomes” (bions).
[2] The organization of protozoa (orgonomia).
.






